
Теперь уже трудно понять, почему так не повезло крупным ископаемым рептилиям. что все, связанное с их жизнью и смертью, стало объектом мифотворчества — от совсем ненаучного до околонаучного и даже научного. Один из таких мифов — утверждение, что динозавры были «теплокровными».
Квалифицированно рассмотреть эту гипотезу могли специалисты по энергетике позвоночных, но в свое время они не обратили на нее внимания из-за ее самоочевидной для них нелепости, позднее аргументированно ответили [16, II. 23]. но. по-видимому, в трудной для восприятия зоологами и палеонтологами форме. Поскольку с точки зрения специалистов по энергетике позвоночных проблема снята, в дальнейшие дискуссии со сторонниками теплокровных динозавров они не вступают. Это было бы правильно, если бы мы имели дело с обычной научной проблемой. Но за последние десятилетия теплокровные динозавры превратились в миф со все большим числом чрезвычайно активных сторонников, выросло целое поколение палеонтологов, которые в него свято верят и тратят свое время на дальнейшее развитие его деталей.
Окончательно меня заставило написать данную статью то, что я увидел в одном из наших лучших университетов: массу биологов, слушающих цикл лекций гастролирующего «профессора по динозаврам» (лекция 10 — «Сезонные миграции динозавров», лекция 12 — «Насиживание яиц у динозавров»).
Цель настоящей статьи — в достаточно доступной форме дать читателю необходимый набор сведений по энергетике рецептных рептилий, птиц и млекопитающих и приемов их анализа в приложении к крупным животным, чтобы он сам мог решить, могли ли гигантские рептилии быть теплокровными.
Автор прекрасно понимает, что таким действием невозможно разрушить миф, но можно помочь разобраться в проблеме тем, кто поверил в него из-за нехватки специальных сведений.
Для многих биологов «все встанет на свои места», просто если мы вернем энергетическим терминам, которыми произвольно пользуются сторонники теплокровных динозавров, их изначальное научное значение (рис. 1).
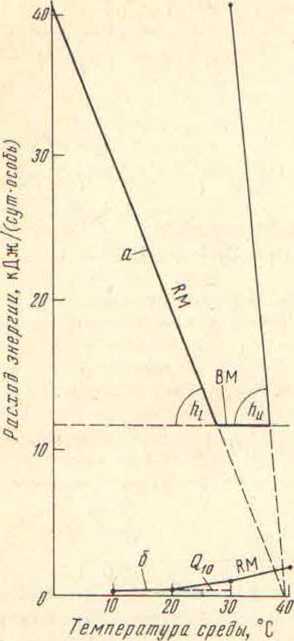
.
У птицы минимальная теплопродукция в покое (ее базальный метаболизм. BMR на порядок выше той максимальной теплопродукции, на которую способна рептилия при температуре среды, равной температуре тела птицы. В ответ на снижение температуры среды птица сначала не меняет теплопродукции, но уменьшает теплоотдачу, плавно переходя от максимальной теплопроводности (hu) покровов к минимальной (hl). При дальнейшем понижении температуры среды птица включает специальную терморегуляционную теплопродукцию (RM = hl * (TB – TA)] компенсирующую возрастающие теплопотери. Этот процесс подчиняется закону охлаждения Ньютона.
Теплокровные животные — это птицы и млекопитающие. Они названы так не столько потому, что у них «теплая кровь» (37—40°С) и такая же температура тела, сколько потому, что они эндотермные организмы, обеспечивающие высокую температуру тела благодаря собственной теплопродукции, мощному потоку тепла, образующегося в организме специально для поддержания температуры тела на высоком уровне. Они называются гомойотермными животными, потому что удерживают температуру тела на почти постоянном уровне, не зависящем от температуры окружающей среды. На снижение внешней температуры, требующее повышения теплоотдачи организма в окружающую среду, они отвечают соответствующим увеличением теплопродукции.
В терморегуляционном ответе участвуют почти все органы и ткани, так как они обладают особой функцией — несократительным термогенезом. Огромная роль в терморегуляционном ответе отведена скелетной мускулатуре, которая характеризуется особой способностью — даже не производя внешней работы увеличивать теплопродукцию за счет терморегуляционного тонуса мышц. Все перечисленное выше называется химической терморегуляцией. Помимо нее гомойотермные животные используют и свои способности к физической терморегуляции — в первую очередь изменяя теплопроводность своих покровов и охлаждаясь за счет кожного и респирационного испарения воды, а также греясь в лучах солнца, от нагретых предметов или, наоборот, прячась от них и т.п.
Холоднокровные (пойкилотермные, эктотермные) — все остальные животные, кроме птиц и млекопитающих. Они названы так не потому, что температура их крови и тела никогда не бывает высокой, а потому, что она обеспечивается не собственной теплопродукцией, а задается окружающей средой (эктотермия) и. следовательно, может быть непостоянной (пойкилотермия). Собственная теплопродукция эктотермного животного слагается из теплопродукции поддерживающих жизнь процессов и теплопродукции работающих мышц, в ней нет специального терморегуляционного компонента. Весь комплекс химической терморегуляции у эктотермных отсутствует. В ответ на снижение температуры среды они снижают уровень своего метаболизма без какого-либо терморегуляционного ответа.
Их скелетная мускулатура может увеличивать теплопродукцию только сокращаясь, когда она производит внешнюю работу, и не способна к терморегуляционному тонусу. Уровень их метаболизма много ниже, чем у эндотермных животных, и образующееся в теле тепло, как правило, успевает рассеиваться без заметного повышения температуры тела, хотя последнее не исключено.
Многие эктотермные животные (насекомые, рептилии) обладают прекрасно развитой физической терморегуляцией. Так, они могут при высоких температурах среды снижать температуру тела за счет кожного и респирационного испарения воды, уходить в тень, воду, контактировать с холодными предметами и т.п. При необходимости они могут повышать температуру тела, очень эффективно абсорбируя солнечные лучи, контактируя с нагретыми предметами и т.п. Тело животного характеризуется определенной теплоемкостью, поэтому остывает не сразу, т.е. обладает тепловой инерцией, что позволяет эктотермному животному переместиться в более холодное место и действовать там, сохраняя некоторое время уровень метаболизма, соответствующий прежнему теплому месту (и наоборот).
У эктотермных животных с развитым терморегуляционным поведением температура тела почти никогда не равна метеорологической температуре того места, где они обитают. Более того, у двух видов, живущих в одном и том же месте, она может быть разной в зависимости от их термофилии.
Некоторые современные рептилии значительную часть времени имеют температуру тела, близкую к температуре тела эндотермных животных. Правильно ли было бы назвать их «теплокровными»? Конечно, нет. Теперь представим себе стабильный климат, позволяющий рептилии всегда достигать температуры тела эндотермных животных, и рептилию огромной массы, с большой тепловой инерцией. Такая рептилия могла иметь всю жизнь почти постоянную высокую температуру тела, но называть на этом основании ее теплокровной или гомойотермной нельзя, так как по физиологическому устройству она остается эктотермным животным.
Как возникло предположение о теплокровности динозавров
Палеонтолог Ромер [49] предположил, что огромный гребень на спине пермских диметродонтов мог выполнять функцию теплообмена. Сходное мнение высказал Родбард [48]. У обоих авторов речь шла не об эндотермности громадных рептилий, а о том, что выросты на теле могли служить для физической терморегуляции, т.е. переноса с помощью кровотока тепла от поверхности тела внутрь его и обратно. Такие приспособления есть у современных рептилий и играют значительную роль в их жизни [12, 17, 28, 54], а у некоторых видов они чрезвычайно эффективны и позволяют на некоторое время получать температуру тела выше или ниже температуры окружающей среды [20]. Это эктотермия. В отличие от нее у эндотермных птиц и млекопитающих более высокая температура тела по сравнению с окружающей средой обеспечивается уровнем теплопродукции организма.
 Неточно поняв Ромера, Стрельников [10] заявил, что динозавры были теплокровными, и нашел объяснение этому в огромном отношении объема их тела к поверхности. Затем палеонтолог Рассел [51] предположил, что кровеносная система динозавров более походила на кровеносную систему эндотермных птиц, чем эктотермных крокодилов, и поэтому температура их тела могла быть высокой. Однако высокая температура тела — это еще не эндотермность. Так, Спотила, исходя из мезозойского климата и физической модели динозавра, получил для него повышение температуры в середине дня из-за абсорбции солнечных лучей вплоть до 40 °С [53].
Неточно поняв Ромера, Стрельников [10] заявил, что динозавры были теплокровными, и нашел объяснение этому в огромном отношении объема их тела к поверхности. Затем палеонтолог Рассел [51] предположил, что кровеносная система динозавров более походила на кровеносную систему эндотермных птиц, чем эктотермных крокодилов, и поэтому температура их тела могла быть высокой. Однако высокая температура тела — это еще не эндотермность. Так, Спотила, исходя из мезозойского климата и физической модели динозавра, получил для него повышение температуры в середине дня из-за абсорбции солнечных лучей вплоть до 40 °С [53].
Палеонтолог Остром [41] в поисках позвоночных — индикаторов особенностей климата в Мезозое осторожно заметил, что динозавры но своей физиологии и уровню активности могли быть непохожи на современных эктотермных рептилий. Вскоре палеонтолог Беккер [13, 14] выдвинул гипотезу о том. что динозавры были эндотермными животными. Ему сразу же возразили морфолог Федучия [30] и специалисты по энергетике рептилий Беннетт и Далзсл [23]. На страницах журнала «Evolution» Беккер ответил на критику новой статьей [15]. там же были помещены замечания участников дискуссии [21, 29, 31,42].
Гипотеза Беккера [13-16] отталкивается от простого морфологического факта: в отличие от современных рептилий динозавры имели полностью поднятую над субcтратом позу, которая характерна для современных птиц и млекопитающих. Из этого делается вывод, что поддержание такой прямой позы требовало, с одной стороны, большого расхода энергии, а с другой — позволяло динозаврам быть очень активными, добывать пищу в сопоставимых с характерными для птиц и млекопитающих количествах. Фактически Беккер постулирует прямую связь между распрямленной позой и высоким уровнем метаболизма, эндотермией. Сам постулат представляется логическим нонсенсом: второе никак не вытекает из первого и не может быть им доказано, на что сразу же указали другие участники дискуссии. Морфология конечностей динозавров, подпирающих тело снизу, могла возникнуть именно для более экономного поддержания массивного тела. Как раз такая морфология конечностей позволяет современным лошадям или слонам расходовать во сне стоя на ногах почти столько же энергии (или чуть больше), сколько они расходуют во сне лежа 126]. У птиц тоже разница в расходе энергии во сне между позой стоя и позой лежа незначительная [38].
Остаются три косвенных палеонтологических аргумента в пользу предложенной гипотезы: высокая степень васкуляризации костей у динозавров, обнаруженная также у крупных птиц и млекопитающих [47], отпечатки следов динозавров, позволяющие предполагать, что они были хотя бы временами очень активны [42], и малое соотношение хищных особей к растительноядным жертвам [15]. Последнее наблюдается у эндотермных млекопитающих и птиц, а у эктотермных (например, у пауков) это отношение выше. Однако Несов [9] привел убедительные данные, свидетельствующие о том, что в ископаемом материале по динозаврам в самом процессе фоссилизации и последующего сохранения нарушается истинное соотношение хищников и жертв.
Пятое палеонтологическое свидетельство того, что динозавры были очень активны — высокое отношение объема их мозга к размерам тела [50] — опровергается безупречным аллометрическим анализом [36]. Оказалось, что в пределах каждого класса позвоночных животных масса мозга увеличивается с повышением массы тела разных видов пропорционально массе тела в степени 0.67, причем линии регрессии для птиц и млекопитающих проходят выше линии регрессии современных рептилий. При этом данные о мозге динозавров ложатся точно на линию регрессии рептилий и оказываются ниже линий регрессии птиц и млекопитающих.
По проблеме эндотермии динозавров был проведен специальный симпозиум «Холодный взгляд на теплокровных динозавров» [57], который позволил некоторым участникам вволю пофантазировать, придумывая динозаврам экстравагантные экологию, физиологию и поведение, но все же не дал никаких палеонтологических доказательств того, что динозавры были эндотермными животными.
Не было недостатка и в моделировании теплового баланса динозавров. Например, Будыко [11], взяв простую модель физического тела заданной массы и мысленно поместив в него нагреватель произвольной мощности, обнаружил, что его «динозавр» разогревается. Однако для настоящего динозавра это означало бы гибель, ибо модель не позволяла остановить разогрев на каком-то приемлемом уровне. Напротив, Спотила с соавт. [53], представив динозавра в виде цилиндра диаметром 1 м и задав ему теплопродукцию крокодила, показали, что в условиях солнечной инсоляции он может столкнуться с проблемой перегрева, которого можно избежать только при очень низком, рептильном уровне теплопродукции.
Кажется очевидным, что нельзя произвольно придумывать энергетику динозавров, а необходимо воспользоваться аллометрическими уравнениями, описывающими энергетику современных рептилий, и посмотреть, что получится, если приложить их к животным, равным по массе динозаврам [11]. В свое время я сделал именно такую попытку [6] и обнаружил, что динозаврам никак не следовало быть эндотермными, иначе они столкнулись бы с неразрешимыми проблемами. Недавно опубликованы утверждения, что в свете новых данных по энергетике рептилий получается, что динозавры могли быть эндотермными [3,44].
Появилось и еще одно палеонтологическое обстоятельство: остатки меловых динозавров обнаружены на территориях современной Субарктики, причем палеоботаники оценивают климат, существовавший в этих местах, как довольно прохладный. Палеонтологи снова стали рассматривать идею об эндотермкоети динозавров как привлекательную. (Чтобы больше не возвращаться к этому частному вопросу о биологии динозавров на севере, заметим, что гаттерия миллионы лет живет в Новой Зеландии в еще худших условиях: она активна при средней температуре тела 11 °С [13].).

Возрождение идеи о теплокровных динозаврах заставило меня обратиться к новым данным по энергетике рептилий и птиц и вновь рассмотреть их в приложении к динозаврам.
Мы будем сравнивать энергетику животных разных размеров тела и разных классов, используя эмпирические уравнения зависимости какой-либо энергетической характеристики животных (у) от их массы тела (m). Степенные аллометрические уравнения имеют форму
у = amb,
где а и b — константы. Первая из них показывает, чему равна характеристика у при m = 1, а вторая — степенной показатель уравнения (или наклон линии регрессии) — показывает, как изменяется у с изменением m. Графическое изображение регрессий степенных уравнений в логарифмической системе координат дает прямые линии, что очень удобно для наглядных сравнений.
Почему возможны рептилии-гиганты
Как должна изменяться с массой тела мощность животных, чтобы они сохраняли равные возможности (изометричность) при передвижении но земле бегом? Достаточно простые теоретические модели предсказывают, что для этого расходуемая при беге мощность должна расти пропорционально массе тела в степени 0,83 (см. [5, 11J). Современные экспериментальные данные по энергетической цене бега замечательно соответствуют этому заключению, причем в отношении бега как рептилий, так и птиц и млекопитающих |43. 56]: максимальное потребление кислорода у бегущих животных пропорционально массе их тела в степени 0.83.
Еще Броди [26] показал, что существует определенное соотношение между расходом энергии при активной работе и в покое, причем животные разных видов сохраняют равные возможности по активности (изометричны), если это соотношение («метаболический диапазон») у них одинаково. На языке аллометрических уравнений принцип Броди будет означать, что для сохранения изометричности необходимо, чтобы были одинаковые степенные показатели в уравнении, описывающем зависимость от массы тела расхода энергии в покое, и в уравнении, описывающем зависимость от массы тела расхода энергии при активности.
Расход энергии в покое (мощность покоя, метаболизм в покое, англ, rest metabolism — RM. кДж/сут) измеряется у спящих животных, завершивших переваривание пищи. У эктотермных животных RM возрастает с увеличением температуры среды, а у эндотермных, наоборот, снижается, пока не достигнет минимального уровня, называемого базальным метаболизмом (англ, basal metabolism — ВМ, кДж/сут). Расходэнергии при активности (мощность активности, метаболизм активности, англ, active metabolism — AM, кДж/сут) измеряется при заданном уровне локомоторной нагрузки (обычно на грани образования кислородной задолженности, т.е. при максимальном аэробном метаболизме). AM у эктотермных животных возрастает с увеличением температуры среды, а у эндотермных почти не зависит от нее.
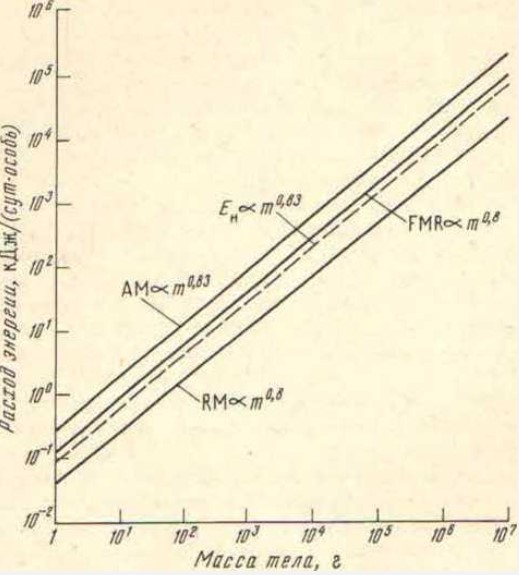
Рис.2. Аллометрическое устройство энергетики класса рептилий (шкалы логарифмические). Изометричность позволяет иметь какие угодно размеры тела. Расход энергии в покое (RM), потребление энергии с нищей (FMR) и предельный расход энергии при беге (AM) изменяются с варьированием массы тела параллельно друг другу, причем именно так, как необходимо для обеспечения равных возможностей при беге по физическим причинам (En).
Если выбрать обычно используемую для сравнения эктотермных животных температуру среды 20 °С, то, согласно последней современной сводке Беннетта [22], у 73 видов рептилий
RM = 0,049 m0,80. (1)
При той же температуре среды максимальный расход аэробной энергии при беге у 18 видов рептилий образует зависимость
AM = 0,311 m0,83. (2)
Следовательно, рептилии расходуют при максимально возможной активности в 6-10 раз больше энергии, чем в покое, причем это соотношение почти не зависит от размеров тела (рис. 2). У рептилий мощность покоя изменяется с массой тела именно таким образом, как этого требует сохранение изометрии при движении. Поэтому для рептилий нет энергетического запрета на эволюционное увеличение массы тела по очень важному показателю — располагаемой (т.е. имеющейся в распоряжении) для движения мощности. Мелкие и гигантские рептилии могут сосуществовать «на равных», без потери конкурентоспособности.
Обратимся теперь к другому важному энергетическому показателю — суточному расходу энергии при существовании в естественных условиях (англ, free metabolic rate — FMR, кДж/сут). Данный показатель расхода энергии в равной степени характеризует и приход ее, т.е. то количество энергии, которое животное потребляет с пищей. Измерения методом дваждытяжелой воды показали [28], что у рептилий летом
FMR = 0,224 m0 80. (3)
И этот признак имеет степенной показатель 0,8, т.е. он изменяется с массой тела параллельно двум предыдущим. Отсюда вытекают два следствия. Первое: можно рассматривать FMR как сумму суточных расходов энергии в покое и при разных уровнях активности. Причем доля расходуемой на активную работу энергии не изменяется с изменением массы тела, а это значит, что крупные рептилии проводят в активном состоянии столько же времени, сколько мелкие. И второе: если рассматривать FMR как суточное потребление энергии пищи, то оказывается, что у рептилий оно зависит от массы тела именно таким образом, как необходимо для обеспечения изометрии по активности. Следовательно, и со стороны потребления пищи у рептилий нет аллометрического запрета на эволюционное увеличение массы тела.
Энергетически рептилии так устроены, что эволюция в направлении гигантизма для них не запрещена. Гигантская рептилия будет обеспечивать себя пищевой энергией и будет снабжена мускульной мощностью ничуть не хуже, чем маленькая. Они могут быть равноактивны по времени, сохранять конкурентоспособность в мире рептилий. Такое выгодное аллометрическое устройство класса возможно для рептилий именно потому, что они не эндотермные животные, для них нет проблемы, как рассеивать избыточное тепло, образующееся в организме. На рис. 2 нанесена еще одна линия (Ен) — необходимая для движения мощность согласно физическим законам (см. [5, 11]). Она параллельна всем трем эмпирическим характеристикам энергетики рептилий.
Чтобы лучше понять, что такое аллометрические запреты на увеличение размеров тела, обратимся к эндотермным птицам и млекопитающим, у которых нет аллометрического соответствия между метаболизмом покоя (базальным метаболизмом, англ, basal metabolism — ВМ), необходимым для бега расходом энергии (Ен). суточным потреблением (и расходом) энергии при существовании в естественных условиях (FMR) и удельной теплоотдачей организма (h) (рис. 3).
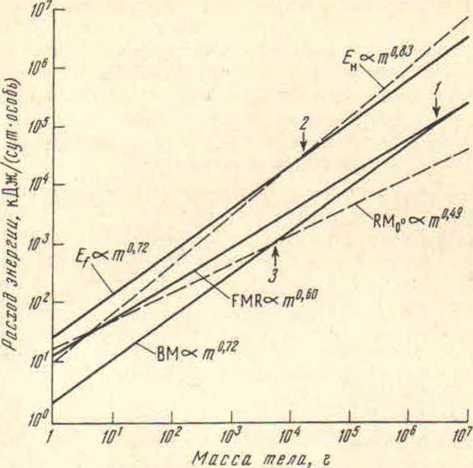
Аллометрическое устройство энергетики у птиц не может быть изометрично, так как оно вынуждено выбирать компромиссные решения между давлениями двух физических требований: необходимая для движения мощность должна расти пропорционально массе в степени 0,83 (линия Eн — необходимая для полета мощность), а теплоотдача может расти только пропорционально массе в степени 0,5 (линия RM, отражающая теплоотдачу в покое при температуре среды 0 °С). Расход энергии при базальном метаболизме (ВМ) не параллелен получению энергии из пищи (FMR), рост располагаемой для полета мощности (Еf) недостаточно крут и т.д. Предельные размеры класса ограничены (вертикальные стрелки): 1 — в FMR не осталось мощности для активности. 2 — располагаемая для полета мощность становится ниже необходимой. 3 — чтобы рассеять теплопродукцию ВМ. нужно сдвинуть нижнюю границу термонейтральной зоны на 40 ниже температуры тела.
Как у всех бегающих животных, у птиц и млекопитающих необходимая для бега мощность (Ен) возрастает, как масса в степени 0,83 [43, 56]. Однако базальный метаболизм, который обеспечивает основу активной мощности, связан с массой тела менее крутой зависимостью: у млекопитающих ВМ пропорционален массе в степени 0,75 [49], а у птиц — в степени 0,72 [40]. «Вилка» между степенным показателем 0,83, с одной стороны, и степенными показателями 0,75 и 0,72, с другой, приводит к тому, что птицы и млекопитающие, для того чтобы сохранять равные возможности в беге по мере увеличения размеров тела, должны расходовать все большее количество энергии по сравнению с их базальным метаболизмом. То число раз, в которое базальный уровень может быть увеличен (метаболический диапазон), ограничено и почти не зависит от массы тела. Поэтому должны существовать предельные размеры тела, при которых движение уже не будет обеспечиваться необходимой для него энергией. В отношении активного полета птиц подобное ограничение особенно наглядно: они не могут на длительное время поднимать метаболизм выше уровня 12 ВМ и этот уровень оказывается ниже требуемого для полета уровня при массе тела более 12 кг. Летающих активным полетом птиц выше этой массы не существует [4].
Еще менее крутой наклон у линий регрессий зависимости энергии самоподдержания в естественной обстановке (FMR) от массы тела: у птиц этот показатель пропорционален массе в степени 0,605 [59], а у млекопитающих — массе в степени 0,67 [38]. «Вилка» между степенными показателями уравнения, описывающего зависимость от массы энергии самоноддержания и уравнения зависимости от массы расхода энергии при активности приводит к тому, что у эндотермных животных время, проводимое в состоянии активности, закономерно падает с увеличением массы тела [59] и при какой-то критической массе должно достичь недопустимо малой величины.
Мелкие и крупные эндотермные животные в отличие от эктотермных рептилий не изометричны между собой, и проблемы, связанные с отклонением от изометрии, возрастают по мере приближения размеров тела к нижнему или верхнему пределам. Этот недостаток «аллометрического устройства» эндотермных животных обусловлен тем. что им приходится сохранять баланс между выработкой тепла в организме и теплоотдачей, рассеиванием этого тепла. Согласно всем современным экспериментальным данным, удельная (на 1°С) теплоотдача растет пропорционально массе тела в степени 0.5 как у млекопитающих [25, 55], так и у птиц [34, 37]. Почему теплоотдача связана с массой не пропорционально поверхности тела, как обычно предполагают, а менее круто, объясняется довольно просто физической моделью, учитывающей неизбежное при росте массы тела увеличение толщины теплоизолирующих покровов [7, 111.
Из-за «вилки» между необходимой для изометрии при движении мощностью (пропорциональной массе в степени 0,83) и теплоотдачей (пропорциональной массе в степени 0,5) зависимости ВМ и FMR от массы у эндотермных животных приобрели промежуточные степенные показатели (0,6 — 0,75), что позволило отодвинуть ограничители крупных размеров, но не отменить их. Поэтому крупные эндотермные животные сталкиваются сразу с двумя проблемами: нехваткой энергии для движения и избытком продуцируемого тепла, который угрожает перегревом тела.
У рептилий с их низкой теплопродукцией не возникает проблемы рассеивания тепла и именно такое их «аллометрическое устройство» изометрично и по-своему совершенно. Если бы крупные динозавры попытались повысить уровень своего метаболизма. они бы сразу же вышли из этого аллометрического устройства и попали в зону запретов, для гигантов непреодолимую. Палеонтологи — сторонники высокого метаболизма динозавров должны осознать, что само существование гигантских рептилий разрешено природой только при условии их полного подобия современным рептилиям как но уровню метаболизма, так и по его зависимости от массы тела.
Метаболизм рептилии при температуре тела эндотермных животных
Многие морфологи и палеонтологи не чувствуют (а потому и забывают) масштаб различия по уровню теплопродукции между эктотермными и эндотермными позвоночными животными. Некоторым кажется, что все дело в температуре тела, что если бы температура тела эктотермного животного поднялась до уровня температуры тела эндотермных животных (37°С у млекопитающих и 40°С у птиц), метаболизм такого животного достиг бы уровня, свойственного эндотермным (т.е. RM рептилий стал бы равным по величине ВМ птиц и млекопитающих). В действительности это совсем не так.
Минимальный (базальный. ВМ, кДж/сут) уровень метаболизма в покое у млекопитающих зависит от массы тела следующим образом (39]:
ВМ = 1,65 m0,75 (4)
У неворобьиных птиц соответствующая зависимость имеет близкие параметры [40]:
ВМ = 2,2 m0,723. (5)
Основываясь на измерениях метаболизма в состоянии покоя, выполненных на рептилиях, температура тела и среды которых была близка к температуре тела эндотермных животных. Беннетт [22] экстраполировал эти величины на температуру тела 40°С и по ним рассчитал зависимость RM при 40°С от массы тела у рептилий:
RM = 0,273 m0,79. (6)
Если по этим уравнениям найти величины ВМ и RM для равных масс тела, то оказывается, что при массе тела 10 г теплопродукция у рептилий в 5,5 раза ниже, чем у млекопитающих, и в 6,9 раза ниже, чем у птиц; при массе тела 1 кг она соответственно ниже в 4,6 и 5,0 раза, а при массе тела 1000 кг в 3,5 и 3.2 раза ниже. Эндотермные животные, следовательно, обладают в несколько раз более высокой теплопродукцией в покое, чем эктотермные при температуре тела эндотермных.
Поэтому неверно представление, что если в Мезозое где-то температура среды была близкой к температуре тела эндотермных животных, то жившие в этих условиях рептилии по уровню метаболизма были близки к современным эндотермным животным. То же относится и к высказываемым предположениям о том, что при разогревании за счет абсорбции солнечной радиации до температуры тела эндотермных динозавры достигали их уровня метаболизма.
Обратимся к максимальной мощности аэробного метаболизма при активности (беге) у рептилий. Беннетт [22] экстраполировал на температуру тела 40°С данные о верхнем предельном расходе энергии рептилиями, полученные при несколько меньших темпера турах, и рассчитал по ним следующее уравнение для AM при 40°С:
AM = 2,83 m0,64. (7)
Согласно этому уравнению, разогретые до 40°С рептилии кратковременно могут при максимально возможной для них активности приближаться по теплопродукции к уровню базального метаболизма эндотермных животных: при массе тела 10 г они достигают уровня ВМ птиц и в 1,3 раза превышают уровень ВМ млекопитающих; при массе тела 1 кг их AM составляет 0,7 от уровня ВМ птиц и 0,8 от уровня ВМ млекопитающих; при массе тела 100 кг эти значения равны соответственно 0,49 и 0.48, а при массе 1000 кг — 0,41 и 0,38.
Сравним теплопродукцию «разогретых» до 40°С рептилий при максимальной активности с теплопродукцией млекопитающих в состоянии бега (AM). Последняя описывается эмпирическим уравнением [45]:
AM = 12,6 m0,73. (8)
Млекопитающие превосходят разогретых до 40°С рептилий по мощности, затрачиваемой при беге, в 5.5 раза при массе тела 10 г. в 8,3 раза при массе тела 1 кг и в 15,4 раза при массе тела 1000 кг.
Неизвестно, какова энергия самоподдержания в естественных условиях (FMR) у рептилий при 40°С. Но известно [22J. что в естественных условиях при более низких температурах тела и среды FMR в 2,0 — 2,5 раза выше, чем расход энергии в покое (RM). Если принять, что и при 40°С FMR равен 2,25 RM. то, умножив на это значение правую часть уравнения (6), можно представить, каковы могли бы быть пищевые потребности рептилий, разогретых до 40°С:
FMR=0,61 m0,79. (9)
У млекопитающих FMR описывается уравнением [38]:
FMR = 7.4 m,67. » (10)
а у птиц уравнением [59J:
FMR = 13,05 m0,605 (11)
Ожидаемый FMR у «разогретых» до 40° рептилий оказывается при массе тела 10 — 100 г ниже, чем у птиц и млекопитающих, в 9 — 13 раз; при массе тела 1000 кг он в 1,7 — 2.3 раза ниже.
В целом можно заключить, что рептилия, «разогретая» до температуры тела эндогермных позвоночных, отнюдь не достигает характерных для них уровней метаболизма (рис. 4). В состоянии максимальной активности она кратковременно приближалась бы к тому уровню минимальной теплопродукции в покое, который имеют птицы и млекопитающие. А уровень метаболизма при активности, свойственный эндотермным животным, был бы в 5-10 раз выше доступного рептилиям уровня. Пищевые потребности рептилии тоже оставались бы на порядок ниже, чем у эндотермных животных.
Предельные возможности теплопродукции эндотермных животных для рептилий совершенно недостижимы (а без использования этих возможностей эндотермия не может сохраняться сколько-нибудь долго). Так. если теплопродукции базального метаболизма оказывается недостаточно для сохранения теплового баланса в покое, птицы и млекопитающие прибавляют к ней теплопродукцию нссократительного термогенеза мускулатуры и ее терморегуляционного тонуса. Максимальная теплопродукция неподвижных животных (Нmax, кДж/сут) при этом достигает следующего уровня [27]:
Hmax=19,1m0,65. (12)
Еще выше у птиц уровень теплопродукции (полный расход энергии) в полете (Ef, кДж/сут) [24]:
Ef= 29,2 m0,72. (13)
Итак, утверждение, что из-за высокой температуры среды в Мезозое динозавры в силу высокой температуры тела были подобны эндотермным животным но своей активности, энергетическим и пищевым потребностям, основано на недоразумении. По всем признакам они должны были оставаться почти на порядок величин ниже эндотермных животных. При этом неважно, за счет чего они достигали близкой к эндотермным животным температуры тела: высокой температуры среды, разогрева при поглощении солнечной инсоляции или даже консервации собственной теплопродукции.

Не только и не столько сама высокая температура тела делает птиц и млекопитающих эндотермными животными, сколько изначальное десятикратное по сравнению с эктотермными животными превышение интенсивности базальных метаболических процессов. Подобные ароморфные преобразования могли происходить только у небольших по размерам рептилий, для гигантов же они бы были смерти подобны.
Ароморфоз, приведший к эндотермии предков птиц и млекопитающих, происходил не только и не столько на уровне морфологических структур (скелет, покровы, сердце, легкие и т.п.), сколько на молекулярном и клеточном уровне. Так, изолированные клетки эндотермных животных потребляют в 10 раз больше кислорода, чем клетки эктотермных при температуре среды 37°С. И более того: клетки, взятые от животных разных размеров (от мыши до лошади), сохраняют аллометрическую зависимость потребления кислорода, свойственную целому животному: интенсивность их дыхания пропорциональна массе тела в степени 0,82 [46]. Иными словами, имеются две параллельные зависимости от массы тела — одна для эктотермных животных, а другая, проходящая на порядок более высоком уровне, — для эндотермных.
«Гигантотермия» у рептилий
Какое бы малое количество тепла ни образовывалось в теле пойкилотермного животного с низким уровнем метаболизма, для его отдачи в среду должен возникать положительный градиент температур между местом, где образуется тепло, и поверхностью тела, где оно рассеивается. Обычно градиент неуловимо мал, поэтому игнорируется. Но в некоторых случаях, например, у насекомых в полете, он достигает достаточно большой величины и даже используется организмом для поддержания постоянной температуры тела. Это явление получило название «факультативная эндотермия» [18]. Изучая морских черепах. Паладино с соавт. [44] установили, что у них во время быстрого плавания образуется градиент температур между внутренними частями тела и средой.
Однако в отличие от насекомых черепахи градиентом никак не управляют и не используют его. Тем не менее авторы дали этому явлению особое название — «гигантотермия» и сделали далеко идущие выводы о том. что гигантские динозавры имели в силу своих огромных размеров повышенную температуру тела. Данная публикация была понята как доказательство эндотермноети динозавров. (Читателю из сказанного ранее должно быть ясно, что высокая температура тела сама по себе еще не делает рептилию по уровню метаболизма равной эндотермным животным.)
Горшков [3] обнаружил, что линии регрессии метаболизма рептилий и удельной минимальной теплоотдачи птиц и млекопитающих пересекаются при какой-то массе тела. На основании этого он заключил, что все рептилии выше определенной критической массы могли быть и были «теплокровными». Поскольку такой же прием когда-то применил я для доказательства обратного [6]. придется остановиться на нем более подробно и с учетом появившихся за эти годы новых обобщающих аллометрических уравнений.
Когда мы хотим понять соотношение между теплопродукцией эндотермного животного и его теплоотдачей, мы традиционно, со времен Шоландера [52] пользуемся законом охлаждения физического тела Ньютона. Согласно этому простому закону теплоотдача нагретого тела (H) прямо пропорциональна его удельной теплоотдаче (h), массе (m) и разности температур тела (Тв) и среды (ТА):
Н = h*m*(TB-TA). (14)
Как видно из этого уравнения, сама его форма предполагает обязательное наличие положительной разности температур между телом и средой. Без этой разности модель бессмысленна, она не работает.
Все аллометрические обобщения экспериментальных данных в отношении удельной теплоотдачи (h) эндотермных животных рассчитываются по участку «терморегуляционной кривой», лежащему ниже термонейтральной зоны, на котором присутствует прирост теплопродукции в ответ на понижение температуры среды, вызванный стремлением эндотермного животного сохранить температуру своего тела постоянной.
Рассчитанный с помощью такого приема показатель (h) отражает минимальную удельную [в кДж/(организм * °С)] теплоотдачу живого тела эндотермного животного. Он весьма физиологичен, но не очень физичен, так как фактически «читает» закон Ньютона в обратном направлении. Тем не менее мы не имеем никаких других надежных аллометрических обобщений в отношении минимальной удельной теплопроводности животных и поэтому вынуждены использовать их и для оценки удельной теплоотдачи эктотермных животных.
Согласно аллометрическим обобщениям удельная теплоотдача защищенного теплоизолирующими покровами животного [в кДж/(сут*особь*°С)] составляет у млекопитающих [55]:
h=0.51m0,51, (15)
а у птиц [40]:
h=0,40m0,49 (16)
У лишенных теплозащитных покровов животных удельная теплоотдача выше. У насекомых она равна [18]
h = 0,71m0,531 (17)
Этот показатель измерен на живых, одетых в перья птицах. Оказалось, что такие птицы способны, когда нужно, рассеивать в 2 раза больше тепла по сравнению с птицами, лишенными перьев (см. уравнение 18).
Подобными механизмами физической регуляции теплоотдачи в достаточной мере обладают и рептилии. Так, морская игуана (масса тела 650 г), включив систему переноса тепла от нагретого камня внутрь, повышает температуру своего тела от 20 до 48°С за 48 мин, а охлаждается на те же 20° за 1,5 ч. Это достигается включением механизма переноса тепла кровообращением то в одном направлении, то в противоположном [20]. Таким образом, рептилии обеспечены всеми возможностями для того, чтобы избавиться от избытка эндогенного тепла, даже если бы он и возникал в каких- либо ситуациях.
Разогрев тела и термостатирование
В живой природе безраздельно господствует закон Вант-Гоффа: скорость химических реакций (Q10) увеличивается в 2 раза с повышением температуры на каждые 10°С. Все без исключения пойкилотермные организмы подчиняются этому закону, причем зачастую Q10 у них даже несколько выше 2.0. Это справедливо и для рептилий [22]. Пока теплопродукция метаболизма успевает рассеиваться без существенного повышения температуры тела, подчинение закону Вант-Гоффа безопасно.
Но если бы метаболизм пойкилотермного животного устойчиво превысил уровень его теплоотдачи, оно оказалось бы (в отличие от простого физического тела) в ловушке саморазогрева: повышенная температура тела ускорила бы метаболизм (в том числе и его экзотермические химические реакции), избыток тепла стал бы еще больше, он бы вновь повысил температуру тела, та в свою очередь вновь вызвала бы увеличение скорости химических реакций и т.д. В конце концов неуправляемый рост температуры тела привел бы ее к критической отметке — температуре денатурации белков и животное бы погибло.
Именно из-за этой опасности гомойотермные животные не пользуются при управлении своим метаболизмом законом Вант-Гоффа. изменяя одновременно и температуру тела и метаболизм (что чисто теоретически возможно и даже перспективнее), а удерживают температуру тела на почти постоянном уровне. Для удержания ее на постоянном уровне они опираются на другой закон — Ньютона, попеременно включая и выключая то нагреватель, то охладитель.
Лишь немногие виды птиц и млекопитающих «решаются» следовать закону Вант- Гоффа сначала для снижения температуры тела в неактивной фазе суточного цикла, а затем для повышения ее перед активной фазой [33]. Причем животные с обратимой гипотермией (колибри, например) все мелкие, массой тела до 10 г. Некоторые насекомые применяют во многом сходный прием — факультативную эндотермию, разогреваясь перед полетом и остывая после него [19]. У мелких животных из-за малых размеров теплоемкость тела (создающая его тепловую инерцию) невелика по сравнению с мощностью их теплопродукции и возможностью теплоотдачи, поэтому они могут управлять процессами разогрева или снижения температуры аккуратно и быстро, не очень рискуя попасть в неуправляемый рост метаболизма и температуры тела. Более крупные птицы и млекопитающие из-за большей тепловой инерции тела «не решаются» использовать преимущества обратимой гипотермии.
Скорость разогрева (t1, °С/мин) одинакова у птиц, млекопитающих и насекомых: она образует единую зависимость от массы тела [32]:
t1= 3,22*m0,51 (23)
При удельной теплоемкости тела животных 3,36 Дж/(г*°С) удельная избыточная (т.е. сверх теплоотдачи) теплопродукция (q, кДж/сут) при разогреве равна:
q= 15,6*m0,51, (24)
а избыточная теплопродукция организма (Q, кДж/сут) соответственно
Q= 15,6*m0,51 (25)
При сравнении ее с максимальной возможной теплопродукцией рептилий (уравнения 3, 6, 7, 9, 19) видно, что она в десятки раз превосходит их возможности.
Из этих уравнений можно рассчитать и время (t2, мин), необходимое для разогрева на 10°С (от 26 до 36°):
t2= 3,1*m0,49. (26)
Время разогрева увеличивается с размерами тела: при массе 1 г оно равно 3,1 мин, 1 кг — 1,5 ч, 100 кг — 14,5 ч, 1 т — 45 ч и 10 т — 139 ч. Даже если бы гипотетические динозавры были способны развивать столь высокий метаболизм, они могли бы за такой длительный промежуток времени потерять контроль над температурой тела. Подобные расчеты можно проделать и в отношении обратного процесса — остывания. На это также требуются десятки часов.
При такой огромной тепловой инерции у гигантов никогда бы не начался процесс выработки термостатирования, переход на пользование законом Ньютона. Они неизбежно оставались бы во власти закона Вант-Гоффа. Поэтому, даже если у них и наблюдались проявления «гигантотермии» (задержки тепла и слабого повышения температуры тела), они никак не могли в процессе эволюции их усиливать, ибо, встав на этот путь, они сразу столкнулись бы с неконтролируемым разогревом.
Эндотермия у рептилий
Единственным известным случаем, когда рептилия произвольно и с ясной целью проявляет эндотермию, остается инкубирующая кладку яиц самка питона [22. 35, 581. В обычных условиях самка обвивает кладку своим телом и просто охраняет ее. Уровень метаболизма самки остается при этом равным величине ее метаболизма в покое при окружающей температуре среды. Но если температура среды становится ниже оптимальной для развития зародышей, самка путем спазматических сокращений мускулатуры повышает температуру тела на 5 — 6°С. При этом потребление кислорода самкой возрастает до предельного для рептилий уровня — примерно в 10 раз по сравнению с уровнем в покое — и приближается к уровню ВМ эндотермных позвоночных. Такой уровень метаболизма питон нс может поддерживать долго и поэтому вскоре прекращает сокращение мышц, вслед за чем температура его тела снижается до температуры окружающей среды.
Инкубирующий питон достигает эндотермного эффекта исключительно за счет сократительного термогенеза мышц. Инкубирующая кладку птица, оставаясь неподвижной, при понижении температуры среды тоже способна увеличить поток тепла в несколько раз, но обеспечивается это иным путем — повышением уровня несократительного термогенеза и терморегуляционного тонуса мышц [8].
Некоторые ученые полагают, что питону не хватает уровня метаболизма, и если бы последний увеличился, то он бы стал настоящим эндотермным животным. Для этого, однако, питону еще необходимы несократительный термогенез почти всех тканей и терморегуляционный тонус скелетных мышц. У птиц и млекопитающих мышца может дознрованно и в разном объеме производить тепло не только сокращаясь для совершения внешней работы, но и когда в этой работе нет необходимости. Благодаря этому возможно изменение теплопродукции независимо от характера деятельности животного, в том числе и в покое. Не обладая терморегуляционным тонусом, питон может продуцировать избыток тепла, пока ничего другого не делает (свернулся и сокращает мышцы). Если же он начнет заниматься чем-то другим, его мышцы будут заняты именно этим и будут продуцировать столько тепла, сколько требуется для данной формы активности.
Нет никаких доказательств того, что динозавры «открыли» терморегуляционный тонус мышц, а без него переход от следования закону Ванг-Г’оффа к использованию закона Ньютона невозможен.
Заключение
У гомойотермных птиц и млекопитающих гигантизм запрещен аллометрическим устройством их энергетики, требующей сохранения теплового баланса между теплопродукцией и теплоотдачей. Поэтому гиганты ограничены как располагаемой для активности мощностью, так и опасностью перегрева. Ограничены они и возможностью потребить необходимое количесгво нищи.
Аллометрическое устройство энергетики рептилий, в силу отсутствия необходимости при их низкой теплопродукции сохранять тепловой баланс, не имеет ограничения на размеры тела, в том числе и на гигантизм. Гигантские рептилии, если аллометрия их метаболизма соответствовала современным, были обеспечены энергией для движения и добывания пищи не хуже, чем более мелкие, и не хуже, чем современные рептилии. Это позволяло гигантам открывать много экологических ниш, в том числе и связанных с разнообразным активным поведением.
Однако все преимущества совершенной эктотермной рептильной аллометрии были бы утеряны, если бы крупные рептилии, давно вошедшие но своим размерам в область запретов для эндогермных животных, «попробовали» стать эндотермными, подобно во много раз более мелким предкам млекопитающих и птиц. Выход на уровень метаболизма эндотермных животных означает 10-кратное его увеличение, при котором гигантское животное неизбежно попадает в состояние неконтролируемого саморазогрева и при этом теряет все преимущества эктотермного гигантизма.
Поэтому даже если в некоторых случаях в теле гигантских рептилий возникала ситуация «гигантотермии» — задержки метаболического тепла с повышением температуры тела, — они должны были стремиться от нее избавиться, так как она опасна из- за риска саморазогрева. Механизмы же физической терморегуляции, позволяющей отвести лишнее тепло из организма, у рептилий прекрасно развиты, и применение их помогало выходить из ситуации «гигантотермии» без потери контроля над температурой тела.
В заключение хотелось бы призвать палеонтологов применять к динозаврам ими же когда-то сформулированный принцип актуализма, рассматривать динозавров как эктотермных рептилий, подчиняющихся общей для этого класса особой аллометрии, и в рамках такого ограничения искать объяснение их биологии. Кроме того, следует отметить, что если бы вопреки всем термодинамическим запретам динозавры решили проблему эндотермного гигантизма, то ничто не мешало бы им сохраниться до наших дней, ведь птицы и млекопитающие эту проблему так и не решили, а посему никогда бы нс смогли конкурировать за ниши гигантов.
Вольф Кицес
Успехи современной биологии. 1998. Т.118. №6. (+список литературы)
P.S. Стоит обратить внимание на различия стиля Виктора Рафаэльевича в своей области (биоэнергетика) и в другой, где пробует популяризовывать («этология человека»). В первом случае он скептичен, критичен, не хватается за модные догадки, а испытывает их всем корпусом данных, почему получается интересное приращение знания. Во втором — прямо наоборот, почему и выходит мифология в этологии. Увы, это общее правило — известный специалист в другой области чаще всего поддерживает модные теории или творит новые, вместо проверки их данными — как он бы автоматически поступил в своей сфере


























Post your comments